Боль. Болевая чувствительность
Поверхностные ткани снабжены нервными окончаниями различных афферентных волокон (Дж.Эрлангер , Г.С.Гассер , 1924). Наиболее толстые, миелинизированные Аb-волокна обладают тактильной чувствительностью. Они возбуждаются при неболезненных прикосновениях и при перемещении. Эти окончания могут служить как полимодальные неспецифические болевые рецепторы только при патологических условиях, например, вследствие возрастания их чувствительности (сенсибилизации) медиаторами воспаления. Слабое раздражение полимодальных неспецифических тактильных рецепторов приводит к чувству зуда . Порог их возбудимости понижают гистамин и серотонин (Г.Штюттген , 1981).
Специфическими первичными болевыми рецепторами (ноцирецепторами) служат два других типа нервных окончаний - тонкие миелинизированные Аd-терминали и тонкие немиелинизированные С-волокна, филогенетически более примитивные. Оба эти типа терминалей представлены и в поверхностных тканях, и во внутренних органах. Некоторые участки тела, например, роговица, иннервируются только Аd и С-афферентами. Ноцирецепторы дают чувство боли в ответ на самые разные интенсивные стимулы - механическое воздействие, термический сигнал (обычно, с температурой более 45-47 0 С), раздражающие химикаты, например, кислоты. Ишемия всегда вызывает боль, поскольку провоцирует ацидоз. Мышечный спазм может вызывать раздражение болевых окончаний из-за относительной гипоксии и ишемии, которые он вызывает, а также вследствие прямого механического смещения ноцирецепторов.
По С-волокнам проводится со скоростью 0,5-2 м/сек медленная, протопатическая, а по миелинизированным, быстропроводящим Аd-волокнам, обеспечивающим скорость проведения от 6 до 30 м/сек, - эпикритическая боль. Кроме кожи, где, по данным А.Г.Бухтиярова (1966), насчитывается не менее 100-200 болевых рецепторов на 1 см 2 , слизистых и роговицы, болевыми рецепторами обоих типов обильно снабжены надкостница (в чём убеждается каждый футболист, получающий при подкате удар по передне-внутренней поверхности голени), а также сосудистые стенки, суставы, мозговые синусы и париетальные листки серозных оболочек.
В висцеральных листках этих оболочек и внутренних органах болевых рецепторов гораздо меньше. К тому же, в паренхиме внутренних органов имеются, исключительно, С-волокна протопатической чувствительности, достигающие спинного мозга в составе вегетативных нервов. Поэтому висцеральную боль труднее локализовать, чем поверхностную. Кроме того, локализация висцеральной боли зависит от феномена “отраженных болей”, механизмы которого рассматриваются ниже. Париетальные брюшина, плевра, перикард, капсулы ретроперитонеальных органов и часть брыжейки имеют не только медленные протопатические С-волокна, но и быстрые эпикритические Аd, связанные со спинным мозгом спинальными нервами. Поэтому боль от их раздражения и повреждения намного острее и чётче локализована. Хирурги еще в доанестезиологическую эпоху заметили, что разрезы кишки менее болезненны, чем рассечение пристеночного листка брюшины. Боли при нейрохирургических операциях максимальны в момент рассечения мозговых оболочек, в то же время кора больших полушарий обладает очень незначительной и строго локальной болевой чувствительностью. Вообще, такой распространённый симптом, как головная боль , практически всегда связан с раздражением болевых рецепторов вне самой ткани мозга. Экстракраниальной причиной головной боли могут быть процессы, локализованные в синусах костей головы, спазм цилиарной и других глазных мышц, тоническое напряжение мышц шеи и скальпа. Интракраниальные причины головной боли - это, в первую очередь, раздражение ноцирецепторов мозговых оболочек. При менингите сильнейшие головные боли охватывают всю голову. Весьма серьёзную головную боль вызывает раздражение ноцирецепторов в мозговых синусах и артериях, особенно в бассейне средне-мозговой артерии. Даже незначительные потери цереброспинальной жидкости (около 20 мл) могут спровоцировать головную боль, особенно, в вертикальном положении тела, поскольку плавчесть мозга меняется, и при уменьшении гидравлической подушки раздражаются болевые рецепторы его оболочек. С другой стороны, избыток цереброспинальной жидкости и нарушение ее оттока при гидроцефалии, отек головного мозга, его набухание при внутриклеточной гипергидратации, полнокровие сосудов мозговых оболочек, вызванное цитокинами при инфекциях, локальные объемные процессы - также провоцируют “самую частую жалобу” - головную боль, так как при этом увеличивается механическое воздействие на болевые рецепторы окружающих собственно мозг структур. Обший принцип локализации головных болей таков, что затылочные боли часто отражают раздражение ноцирецепторов сосудов и мозговых оболочек под tentorium, а надпалаточные раздражители и стимуляция верхней поверхности самой палатки проявляются лобно-теменными болями. Знакомая очень значительной части человечества “головная боль с похмелья” имеет комплексный патогенез, включая индуцированное алкоголем полнокровие мозговых оболочек и внутриклеточную гипергидратацию. Патофизиология некоторых форм головной боли, тесно связанных с гуморальными медиаторами болевой и антиболевой систем и с проводниковыми механизмами этих систем, в частности, мигрени, отдельно рассматривается ниже.
Паренхима селезёнки, почки, печени и легкого совершенно лишена ноцирецепторов. Зато ими богато снабжены бронхи, желчевыводящие пути, капсулы и сосуды этих органов. Даже значительные по размеру абсцессы печени или лёгкого могут быть почти безболезненными. Однако, плеврит или холангит порой дают серьёзный болевой синдром, сами по себе не будучи тяжёлыми. Висцеральные болевые рецепторы отличаются ещё и тем, что развивают сравнительно слабый ответ на строго локальное повреждение органа, например, хирургический разрез. Однако, при диффузном вовлечении ткани в альтерацию (на фоне ишемии, при действии литических ферментов и раздражающих химикатов, при спазмах и перерастяжении полых органов), их чувствительность под воздействием медиаторов воспаления стремительно растёт, и от них исходит сильная импульсация.
Болевые рецепторы претендуют на уникальное положение в человеческом теле. Это единственный тип чувствительных рецепторов, который не подлежит какой бы то ни было адаптации или десенсибилизации под воздействием длящегося или повторяющегося сигнала. Ноцирецепторы не повышают при этом порог своей возбудимости, как это делают другие, например, холодовые сенсоры. Следовательно, рецептор не “привыкает” к боли. Более того, в ноцирецептивных нервных окончаниях имеет место прямо противоположное явление - сенсибилизация болевых рецепторов сигналом. При воспалении, повреждениях тканей (особенно, внутренних органов) и при повторных и длительных болевых раздражителях порог возбудимости ноцирецепторов снижается. Даже легчайшие прикосновения к ожоговой поверхности крайне болезненны. Это явление называется первичной гиперальгезией . Пальпация внутренних органов, даже если она интенсивна, не причиняет боли, если нет их воспаления. Однако при воспалении чувствительность молчащих внутренних ноцирецепторов настолько увеличивается, что врач регистрирует болевые симптомы. Поколачивание по области почек, безболезненное в отсутствие их повреждений, ведет к болевому ощущению в случае, если почечные ноцирецепторы сенсибилизированы медиаторами воспаления (положительный симптом Пастернацкого). Легко отметить, что если бы происходила адаптация болевых рецепторов, все хронические деструктивные процессы были бы безболезненны и боль утратила бы свою функцию сигнала, который, по выражению И.П.Павлова , “побуждает отбросить то, что угрожает жизненному процессу”.
Называя болевые сенсоры рецепторами, мы должны подчеркнуть, что применение к ним этого термина носит условный характер - ведь это свободные нервные окончания, лишённые каких бы то ни было специальных рецепторных приспособлений.
Нейрохимические механизмы раздражения ноцирецепторов хорошо изучены. Их основным стимулятором является брадикинин. В ответ на повреждение клеток близ ноцирецептора освобождаются этот медиатор, а также простагландины, лейкотриены и ионы калия и водорода. Простагландины и лейкотриены сенсибилизируют ноцирецепторы к кининам, а калий и водород облегчают их деполяризацию и возникновение в них электрического афферентного болевого сигнала. Возбуждение распространяется не только афферентно, но и антидромно, в соседние ветви терминали. Там оно приводит к секреции вещества Р. Этот нейропептид, о котором уже упоминалось, вызывает вокруг терминали паракринным путём гиперемию, отек, дегрануляцию тучных клеток и тромбоцитов. Освобождаемые при этом гистамин, серотонин, простагландины сенсибилизируют ноцирецепторы, а химаза и триптаза мастоцитов усиливают продукцию их прямого агониста - брадикинина. Следовательно, при повреждении ноцирецепторы действуют и как сенсоры, и как паракринные провокаторы воспаления. Вблизи ноцирецепторов, как правило, располагаются симпатические норадренергические постганглионарные нервные окончания, которые способны модулировать чувствительность ноцирецепторов. При травмах периферических нервов нередко развивается так называемая каузалгия - патологически повышенная чувствительность ноцирецепторов в области, иннервируемой повреждённым нервом, сопровождаемая жгучими болями и даже признаками воспаления без видимых местных повреждений. Механизм каузалгии связан с гипералгизующим действием симпатических нервов, в частности, выделяемого ими норадреналина, на состояние болевых рецепторов. Возможно, при этом происходит секреция вещества Р и других нейропептидов симпатическими нервами, что и обусловливает воспалительные симптомы. Явление каузалгии представляет собой, в полном смысле, нейрогенное воспаление, хотя оно вызывается не нервным, а паракринным способом (см. также выше, о роли нервной регуляции в воспалении).
Как впервые предположили У.Кэннон и А.Розенблют (1951) паракринная безымпульсная нейропептидэргическая деятельность нервных окончаний в тканях и составляет реальную основу явления, которое в течение более чем 100 лет, от Ф. Мажанди (1824) до Л.А. Орбели (1935) и А.Д. Сперанского , (1937), именовали нервной трофикой .
Дата добавления: 2015-05-19 | Просмотры: 985 | Нарушение авторских прав
| | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | |
Болевые рецепторы (ноцирецепторы)
Ноцицепторы - специфические рецепторы, при возбуждении которых возникают болевые ощущения. Это свободные нервные окончания, которые могут быть расположены в любых органах и тканях и связаны с проводниками болевой чувствительности. Эти нервные окончания + проводники болевой чувствительности = сенсорная болевая единица. Большинство ноцицепторов имеет двойной механизм возбуждения, т. е. могут возбуждаться под действием повреждающих и неповреждающих агентов.
Периферический отдел анализатора представлен рецепторами боли, которые по предложению Ч. Шеррингтона называют ноцицепторами (от лат. разрушать). Это высокопороговые рецепторы, реагирующие на разрушающие воздействия.
Болевые рецепторы являются свободными окончаниями чувствительных миелиновых и безмиелиновых нервных волокон, расположенных в коже, слизистых оболочках, надкостнице, зубах, мышцах, органах грудной и брюшной полости и других органах и тканях. Число ноцирецепторов в коже человека примерно 100-200 на 1 кв. см. кожной поверхности. Общее число таких рецепторов достигает 2-4 млн.
По механизму возбуждения ноцицепторы делят на следующие основные виды болевых рецепторов:
- 1. Механоноцицепторы: реагируют на сильные механические раздражители, проводят быструю боль и быстро адаптируются. Механоноцицепторы расположены преимущественно в коже, фасциях, сухожилиях, суставных сумках и слизистых оболочках пищеварительного тракта. Это свободные нервные окончания миелинизированных волокон типа А-дельта со скоростью проведения возбуждения 4 - 30 м/с. Они реагируют на действие агента, вызывающего деформацию и повреждение мембраны рецептора при сжатии или растяжении тканей. Для большинства этих рецепторов характерна быстрая адаптация.
- 2. Хемоноцицепторы расположены также на коже и в слизистых оболочках, но превалируют во внутренних органах, где локализуются в стенках мелких артерий. Они представлены свободными нервными окончаниями немиелинизированных волокон типа С со скоростью проведения возбуждения 0,4 - 2 м/с. Специфическими раздражителями для этих рецепторов являются химические вещества (алгогены), но только те, которые отнимают кислород у тканей, нарушают процессы окисления.
Выделяют три типа алгогенов, каждый из которых имеет собственный механизм активации хемоноцицепторов.
Тканевые алгогены (серотонин, гистамин, ацетилхолин и др.) образуются при разрушении тучных клеток соединительной ткани и, попадая в интерстициальную жидкость, непосредственно активируют свободные нервные окончания.
Плазменные алгогены (брадикинин, каллидин и простагландины), выполняя роль модуляторов, повышают чувствительность хемоноцицепторов к ноцигенным факторам.
Тахикинины выделяются при повреждающих воздействиях из окончаний нервов (к ним относится вещество П - полипептид), они воздействуют местно на мембранные рецепторы того же нервного окончания.
3. Термоноцицепторы: реагируют на сильные механические и тепловые (более 40 градусов) раздражители, проводят быструю механическую и термическую боль, быстро адаптируются.
Оглавление темы "Температурная чувствительность. Висцеральная чувствительность. Зрительная сенсорная система.":1. Температурная чувствительность. Тепловые рецепторы. Холодовые рецепторы. Температурное восприятие.
2. Боль. Болевая чувствительность. Ноцицепторы. Пути болевой чувствительности. Оценка боли. Ворота боли. Опиатные пептиды.
3. Висцеральная чувствительность. Висцерорецепторы. Висцеральные механорецепторы. Висцеральные хеморецепторы. Висцеральная боль.
4. Зрительная сенсорная система. Зрительное восприятие. Проецирование световых лучей на сетчатку глаза. Оптическая система глаза. Рефракция.
5. Аккомодация. Ближайшая точка ясного видения. Диапазон аккомодации. Пресбиопия. Возрастная дальнозоркость.
6. Аномалии рефракции. Эмметропия. Близорукость (миопия). Дальнозоркость (гиперметропия). Астигматизм.
7. Зрачковый рефлекс. Проекция зрительного поля на сетчатку. Бинокулярное зрение. Конвергенция глаз. Дивергенция глаз. Поперечная диспарация. Ретинотопия.
8. Движения глаз. Следящие движения глаз. Быстрые движения глаз. Центральная ямка. Саккадамы.
9. Преобразование энергии света в сетчатке. Функции (задачи) сетчатки. Слепое пятно.
10. Скотопическая система сетчатки (ночное зрение). Фотопическая система сетчатки (дневное зрение). Колбочки и палочки сетчатки. Родопсин.
Боль. Болевая чувствительность. Ноцицепторы. Пути болевой чувствительности. Оценка боли. Ворота боли. Опиатные пептиды.
Боль определяется как неприятное сенсорное и эмоциональное переживание, связанное с истинным или потенциальным повреждением ткани или описываемое в терминах такого повреждения. В отличие от других сенсорных модальностей боль всегда субъективно неприятна и служит не столько источником информации об окружающем мире, сколько сигналом повреждения или болезни. Болевая чувствительность побуждает к прекращению контактов с повреждающими факторами среды.
Болевые рецепторы или ноцицепторы представляют собой свободные нервные окончания, расположенные в коже, слизистых оболочках, мышцах, суставах, надкостнице и во внутренних органах. Чувствительные окончания принадлежат либо безмякотным, либо тонким миелинизиро-ванным волокнам, что определяет скорость проведения сигналов в ЦНС и дает повод к различению ранней боли, короткой и острой, возникающей при проведении импульсов с большей скоростью по миелиновым волокнам, а также поздней, тупой и длительной боли, в случае проведения сигналов по безмякотным волокнам. Ноцицепторы относятся к полимодальным рецепторам, поскольку могут активироваться стимулами разной природы: механическими (удар, порез, укол, щипок), термическими (действие горячих или холодных предметов), химическими (изменение концентрации ионов водорода, действие гистамина, брадикинина и ряда других биологически активных веществ). Порог чувствительности ноцицепторов высок, поэтому лишь достаточно сильные стимулы вызывают возбуждение первичных сенсорных нейронов: например, порог болевой чувствительности для механических стимулов примерно в тысячу раз превышает порог тактильной чувствительности.
Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и образуют синапсы с нейронами второго порядка, расположенными в задних рогах спинного мозга. Аксоны нейронов второго порядка переходят на противоположную сторону спинного мозга, где образуют спиноталамический и спиноретикулярный тракты. Спиноталамический тракт оканчивается на нейронах нижнего заднелате-рального ядра таламуса, где происходит конвергенция проводящих путей болевой и тактильной чувствительности. Нейроны таламуса образуют проекцию на соматосенсорную кору: этот путь обеспечивает осознаваемое восприятие боли, позволяет определять интенсивность стимула и его локализацию.
Волокна спиноретикулярного тракта оканчиваются на нейронах ретикулярной формации, взаимодействующих с медиальными ядрами таламуса. При болевом раздражении нейроны медиальных ядер таламуса оказывают модулирующее влияние на обширные регионы коры и структуры лимбической системы, что приводит к повышению поведенческой активности человека и сопровождается эмоциональными и вегетативными реакциями. Если спиноталамический путь служит для определения сенсорных качеств боли, то спиноретикулярный путь предназначен играть роль сигнала общей тревоги, оказывать на человека общее возбуждающее действие.

Субъективную оценку боли определяет соотношение нейронной активности обоих путей и зависимая от нее активация антиноцицептивных нисходящих путей, способных изменить характер проведения сигналов от ноцицепторов . В сенсорную систему болевой чувствительности встроен эндогенный механизм ее уменьшения путем регуляции порога синаптических переключений в задних рогах спинного мозга («ворота боли »). На передачу возбуждения в этих синапсах влияют нисходящие волокна нейронов серого вещества вокруг водопровода, голубого пятна и некоторых ядер срединного шва. Медиаторы этих нейронов (энкефалин, серотонин, норадреналин) тормозят активность нейронов второго порядка в задних рогах спинного мозга, чем уменьшают проведение афферентных сигналов от ноци-цепторов.
Аналгезирующим (обезболивающим ) действием обладают опиатные пептиды (дайнорфин , эндорфины ), синтезируемые нейронами гипоталамуса, которые имеют длинные отростки, проникающие в другие отделы головного мозга. Опиатные пептиды присоединяются к специфическим рецепторам нейронов лимбической системы и медиальной области таламуса, их образование увеличивается при некоторых эмоциональных состояниях, стрессе, продолжительных физических нагрузках, у беременных женщин незадолго перед родами, а также в результате психотерапевтического воздействия или акупунктуры . В результате повышенного образования опиатных пептидов активируются антиноцицептивные механизмы и повышается порог болевой чувствительности. Баланс между ощущением боли и ее субъективной оценкой устанавливается с помощью лобных областей мозга, участвующих в процессе восприятия болевых стимулов. При поражении лобных долей (например, в результате травмы или опухоли) порог болевой чувствительности не изменяется и поэтому сенсорный компонент болевого восприятия сохраняется неизменным, однако субъективная эмоциональная оценка боли становится иной: она начинает восприниматься лишь как сенсорное ощущение, а не как страдание.
Физиология боли
В узком смысле слова боль – это неприятное ощущение, возникающее при действии сверхсильных раздражителей, вызывающих структурно-функциональные нарушения в организме. Отличия боли от других ощущений в том, что она не информирует мозг о качестве раздражителя, а указывает на то, что раздражитель является повреждающим. Другой особенностью болевой сенсорной системы является наиболее сложной и мощный ее эфферентный контроль.
Болевой анализатор запускает в ЦНС несколько программ ответа организма на боль. Следовательно, боль имеет несколько компонентов. Сенсорный компонент боли характеризует ее как неприятное, тягостное ощущение; аффективный компонент – как сильную отрицательную эмоцию; мотивационный компонент – как отрицательную биологическую потребность, запускающую поведение организма, направленное на выздоровление. Моторный компонент боли представлен различными двигательными реакциями: от безусловных сгибательных рефлексов до двигательных программ антиболевого поведения. Вегетативный компонент характеризует нарушение функций внутренних органов и обмена веществ при хронических болях. Когнитивный компонент связан с самооценкой боли, боль при этом выступает как страдание. При деятельности других систем эти компоненты слабо выражены.
Биологическая роль боли определяется несколькими факторами. Боль исполняет роль сигнала об угрозе или повреждении тканей организма и предупреждает их. Боль имеет познавательную функцию: человек через боль учится избегать возможных опасностей внешней среды. Эмоциональный компонент боли выполняет функцию подкрепления при образовании условных рефлексов. Боль является фактором мобилизации защитно-приспособительных реакций организма при повреждении его тканей и органов.
Выделяют два вида боли – соматическую и висцеральную. Соматическую боль подразделяют на поверхностную и глубокую Поверхностная боль может быть ранняя (быстрая, эпикрическая) и поздняя (медленная, протопатическая).
Существуют три теории боли.
1. Теория интенсивности была предложена Э.Дарвином и А.Гольдштейнером. По этой теории боль не является специфическим чувством и не имеет своих специальных рецепторов. Она возникает при действии сверхсильных раздражителей на рецепторы пяти известных органов чувств. В формировании боли участвуют конвергенция и суммация импульсов в спинном и головном мозге.
2. Теория специфичности была сформулирована немецким физиологом М.Фреем. В соответствии с этой теорией боль является специфическим чувством, имеющим собственный рецепторный аппарат, афферентные волокна и структуры головного мозга, перерабатывающие болевую информацию. Эта теория в дальнейшем получила более полное экспериментальное и клиническое подтверждение.
3. Современная теория боли базируется преимущественно на теории специфичности. Было доказано существование специфичных болевых рецепторов. Вместе с тем в современной теории боли использовано положение о роли центральной суммации и конвергенции в механизмах боли. Наиболее крупными достижениями современной теории боли является разработка механизмов центрального восприятия боли и запуска противоболевой системы организма.
Болевые рецепторы
Болевые рецепторы являются свободными окончаниями чувствительных миелиновых нервных волокон Аδ и немиелиновых волокон С. Они найдены в коже, слизистых оболочках, надкостнице, зубах, мышцах, суставах, внутренних органах и их оболочках, сосудах. Их нет в нервной ткани головного и спинного мозга. Наибольшая их плотность имеется на границе дентина и эмали зуба.
Выделяют следующие основные типы болевых рецепторов:
1. Механоноцицепторы и механотермические ноцицепторы Аδ-волокон реагируют на сильные механические и термические раздражители, проводят быструю механическую и термическую боль, быстро адаптируются; расположены преимущественно в коже, мышцах, суставах, надкостнице; их афферентные нейроны имеют малые рецептивные поля.
2. Полисенсорные ноцицепторы С-волокон реагируют на механические, термические и химические раздражители, проводят позднюю плохо локализованную боль, медленно адаптируются; их афферентные нейроны имеют большие рецептивные поля.
Болевые рецепторы возбуждаются тремя видами раздражителей:
1. Механические раздражители, создающие давление более 40г/мм 2 при сдавливании, растяжении, сгибании, скручивании.
2. Термические раздражители могут быть тепловыми (> 45 0 С) и холодовыми (< 15 0 С).
3. Химические раздражители, освобождающиеся из поврежденных клеток тканей, тучных клеток, тромбоцитов (К + , Н + , серотонин, ацетилхолин, гистамин), плазмы крови (брадикинин, каллидин) и окончаний ноцицептивных нейронов (вещество Р). Одни из них возбуждают ноцицепторы (К + , серотонин, гистамин, брадикинин, АДФ), другие сенсибилизируют их.
Свойства болевых рецепторов: болевые рецепторы имеют высокий порог возбуждения, что обеспечивает их ответ только на чрезвычайные раздражители. Ноцицепторы С-афферентов плохо адаптируются к длительно действующим раздражителям. Возможно повышение чувствительности болевых рецепторов – снижение порога их раздражения при многократной или длительной стимуляции, что называется гипералгезией. При этом ноцицепторы способны отвечать на стимулы субпороговой величины, а также возбуждаться раздражителями других модальностей.
Проводящие пути болевой чувствительности
Нейроны, воспринимающие болевую импульсацию. От болевых рецепторов туловища, шеи и конечностей Аδ- и С-волокна первых чувствительных нейронов (их тела находятся в спинальных ганглиях) идут в составе спинномозговых нервов и входят через задние корешки в спинной мозг, где разветвляются в задних столбах и образуют синаптические связи прямо или через интернейроны со вторыми чувствительными нейронами, длинные аксоны которых входят в состав спиноталамических путей. При этом они возбуждают два вида нейронов: одни нейроны активируются только болевыми стимулами, другие – конвергентные нейроны – возбуждаются также и неболевыми стимулами. Вторые нейроны болевой чувствительности преимущественно входят в состав боковых спиноталамических путей, которые и проводят большую часть болевых импульсов. На уровне спинного мозга аксоны этих нейронов переходят на сторону, противоположную раздражению, в стволе головного мозга они доходят до таламуса и образуют синапсы на нейронах его ядер. Часть болевой импульсации первых афферентных нейронов переключаются через интернейроны на мотонейроны мышц-сгибателей и участвуют в формировании защитных болевых рефлексов. В боковом спиноталамическом пути выделяют эволюционно более молодой неоспиноталамический путь и древний палеоспиноталамический путь.
Неоспиноталамический путь проводит болевые сигналы по Аδ-волокнам преимущественно в специфические сенсорные (вентральные задние) ядра таламуса, имеющие хорошую топографическую проекцию периферии тела. Кроме этого небольшая часть импульсов поступает в ретикулярную формацию ствола и далее в неспецифические ядра таламуса. Передача возбуждения в синапсах этого пути осуществляется с помощью быстродействующего медиатора глутамата. Из специфических ядер таламуса болевые сигналы передаются преимущественно в сенсорную кору больших полушарий. Эти особенности формируют основную функцию неоспиноталамического пути – проведение «быстрой» боли и восприятие ее с высокой степенью локализации.
Палеоспиноталамический путь проводит болевые сигналы по С-волокнам преимущественно в неспецифические ядра таламуса прямо или после переключения в нейронах ретикулярной формации ствола мозга. Передача возбуждения в синапсах этого пути происходит более медленно. Медиатором является вещество Р. Из неспецифических ядер импульсация поступает в сенсорную и другие отделы коры больших полушарий. Небольшая часть импульсации поступает и в специфические ядра таламуса. В основном волокна этого пути оканчиваются на нейронах 1) неспецифических ядер таламуса; 2) ретикулярной формации; 3) центрального серого вещества; 4) голубого пятна; 5) гипоталамуса. Через палеоспиноталамический путь проводится «поздняя», плохо локализуемая боль, формируются аффективно-мотивационные проявления болевой чувствительности.
Кроме этого болевая чувствительность частично проводится по другим восходящим путям: переднему спиноталамическому, тонкому и клиновидному путям.
Вышеназванные пути проводят и другие виды чувствительности: температурную и тактильную.
Роль коры больших полушарий в восприятии боли
Полноценное чувственное восприятие боли организмом без участия коры головного мозга невозможно.
Первичное проекционное поле болевого анализатора находится в соматосенсорной коре задней центральной извилины. Оно обеспечивает восприятие «быстрой» боли и идентификацию места ее возникновения на теле. Для более точной идентификации локализации боли в процесс обязательно включается и нейроны моторной коры передней центральной извилины.
Вторичное проекционное поле расположено в соматосенсорной коре на границе пересечения центральной борозды с верхним краем височной доли. Нейроны данного поля имеют двусторонние связи с ядрами таламуса, что позволяет этому полю избирательно фильтровать, проходящие через таламус возбуждения болевого характера. А это в свою очередь позволяет данному полю вовлекаться в процессы, связанные с извлечением из памяти энграммы необходимого поведенческого акта, его реализации в деятельности эффекторов и оценки качества достигнутого полезного результата. Двигательные компоненты болевого поведения формируются в совместной деятельности моторной и премоторной коры, базальных ганглиев и мозжечка.
Лобная кора играет важную роль в восприятии боли. Она обеспечивает самооценку боли (ее когнитивный компонент) и формирование целенаправленного болевого поведения.
Лимбическая система (поясная извилина, гиппокамп, зубчатая извилина, миндалевидный комплекс височной доли) получает болевую информацию от передних ядер таламуса и формирует эмоциональный компонент боли, запускает вегетативные, соматические и поведенческие реакции, обеспечивающие приспособительные реакции к болевому раздражителю.
Некоторые виды болевых ощущений
Существуют боли, которые названы проекционными или фантомными . Их возникновение основано на законе проекции боли: какая бы часть афферентного пути не раздражалась, боль ощущается в области рецепторов данного сенсорного пути. По современным данным в формировании данного вида болевого ощущения участвуют все отделы болевой сенсорной системы.
Существуют также так называемые отраженные боли: когда боль ощущается не только в пораженном органе, но и в соответствующем дерматоме тела. Участки поверхности тела соответствующего дерматома, где возникает ощущение боли, назвали зонами Захарьина – Геда . Возникновение отраженных болей связано с тем, что нейроны, проводящие болевую импульсацию от рецепторов пораженного органа и кожи соответствующего дерматома, конвергируют на одном и том же нейроне спиноталамического пути. Раздражение этого нейрона с рецепторов пораженного органа в соответствии с законом проекции боли приводит к тому, что боль ощущается и в области кожных рецепторов.
Антиноцицептивная система
Антиболевая система состоит из четырех уровней: спинального, стволового, гипоталамического и коркового.
1. Спинальный уровень антиноцицептовной системы. Важным ее компонентом является «воротный контроль» спинного мозга, концепция которого имеет следующие основные положения: передача болевых нервных импульсов с первых нейронов на нейроны спиноталамических путей (вторые нейроны) в задних столбах спинного мозга модулируется спинальным воротным механизмом – тормозными нейронами, расположенными в желатинозном веществе спинного мозга. На этих нейронах оканчиваются разветвления аксонов различных сенсорных путей. В свою очередь нейроны желатинозной субстанции оказывают пресинаптическое торможение в местах переключения первых и вторых нейронов болевых и других сенсорных путей. Часть нейронов являются конвергентными: на них образуют синапсы нейроны не только от болевых, но и от других рецепторов. Спинальный воротный контроль регулируется соотношением импульсов, поступающих по афферентным волокнам большого диаметра (неболевая чувствительность) и малого диаметра (болевая чувствительность). Интенсивный поток импульсов по волокнам большого диаметра ограничивает передачу болевых сигналов на нейроны спиноталамических путей (закрывает «ворота»). Напротив, интенсивный поток болевых импульсов по первому афферентному нейрону, ингибируя тормозные интернейроны, облегчает передачу болевых сигналов на нейроны спиноталамических путей (открывает «ворота»). Спинальный воротный механизм находится под постоянным влиянием нервных импульсов структур ствола мозга, которые передаются по нисходящим путям как на нейроны желатинозной субстанции, так и на нейроны спиноталамических путей.
2. Стволой уровень антиноцицептивной системы. К стволовым структурам противоболевой системы относятся, во-первых, центральное серое вещество и ядра шва, образующие единый функциональный блок, во-вторых, крупноклеточное и парагигантоклеточное ядра ретикулярной формации и голубое пятно. Первый комплекс блокирует прохождение болевой импульсации на уровне релейных нейронов ядер задних рогов спинного мозга, а также релейных нейронов сенсорных ядер тройничного нерва, образующих восходящие пути болевой чувствительности. Второй комплекс возбуждает почти всю антиноцицептивную систему (см.рис.1).
3. Гипоталамический уровень антиноцицептивной системы, с одной стороны, функционирует самостоятельно, а с другой – выступает как настройка, контролирующая и регулирующая антиноцицептивные механизмы стволового уровня за счет связей гипоталамических нейронов разной ядерной принадлежности и разной нейрохимической специфичности. Среди них идентифицированы нейроны, в окончаниях аксонов которых выделяются энкефалины, β-эндорфин, норадреналин, дофамин см.рис.2).
4. Корковый уровень антиноцицептивной системы. Объединяет и контролирует деятельность антиноцицептивных структур различного уровня соматосенсорная область коры больших полушарий. При этом наиболее важную роль в активации спинальных и стволовых структур играет вторичная сенсорная область. Ее нейроны образуют наибольшее количество волокон нисходящего контроля болевой чувствительности, направляющиеся к задним рогам спинного мозга и ядрам ствола головного мозга. Вторичная сенсорная кора видоизменяет активность стволового комплекса антиноцицептивной системы. Кроме этого соматосенсорные поля коры больших полушарий контролируют проведение афферентных болевых импульсов через таламус. Кроме таламуса, кора большого мозга регламентирует прохождение болевой импульсации в гипоталамусе, лимбической системе, ретикулярной формации, спинном мозге. Ведущая роль в обеспечении кортико-гипоталамических влияний отводится нейронам лобной коры.
Медиаторы антиноцицептивной системы
К медиаторам противоболевой системы относят пептиды, которые образуются в головном мозге, аденогипофизе, мозговом слое надпочечников, желудочно-кишечном тракте, плаценте из неактивных предшественников.. Сейчас к опиатным медиаторам антиноцицептивной системы относят: 1) ά-, β-, γ-эндорфины; 2) энкефалины; 3) динорфины. Эти медиаторы действуют на три вида опиатных рецепторов: μ-, δ-, κ-рецепторы. Наиболее селективным стимулятором μ-рецепторов являются эндорфины, δ-рецепторов – энкефалины, а κ-рецепторов – динорфины. Плотность μ- и κ-рецепторов высокая в коре больших полушарий и в спинном мозгу, средняя – в стволе головного мозга; плотность δ-рецепторов средняя в коре больших полушарий и спинном мозгу, малая – в стволе мозга. Опиоидные пептиды угнетают действие веществ, вызывающих боль, на уровне ноцицепторов, уменьшают возбудимость и проводимость болевой импульсации, угнетают вызванную реакцию нейронов, находящихся в составе цепей, передающих болевую импульсацию. Эти пептиды поступают к нейронам болевой сенсорной системы с кровью и ликвором. Выделяются опиоидные медиаторы в синаптических окончаниях нейронов противоболевой системы. Аналгезирующий эффект эндорфинов высокий в головном и спинном мозге, эффект энкефалинов в этих структурах средний, эффект динорфинов в головном мозге низкий, в спинном мозге – высокий.
Рис.1. Взаимодействие основных элементов обезболивающей системы первого уровня: ствол мозга – спиной мозг. (светлые кружки – возбуждающие нейроны, черные – тормозящие).
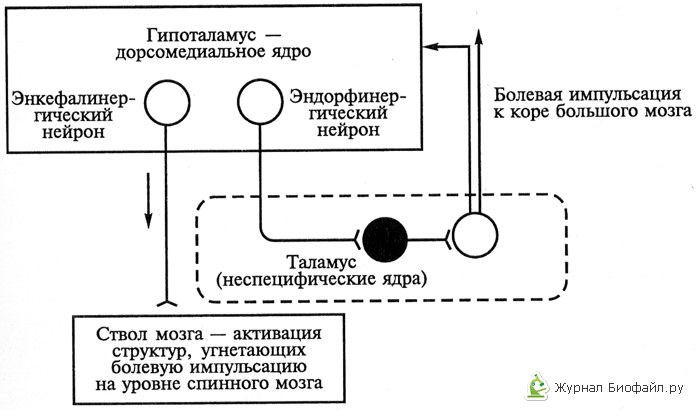
Рис.2. Механизм работы обезболивающей системы организма второго уровня (гипоталамус – таламус – ствол мозга) с помощью опиоидов.
Светлые кружки – возбуждающие нейроны, черные - тормозящие.
Степень выраженности болевого ощущения не определяется одной лишь силой экзогенного или эндогенного болевого воздействия. Во многом оно зависит от соотношения активностей ноцицептивного и антиноцицептивного отделов системы боли, что имеет приспособительное значение.
Поверхностные ткани снабжены нервными окончаниями различных афферентных волокон. Наиболее толстые, миелинизированные Аβ-волокна обладают тактильной чувствительностью. Они возбуждаются при неболезненных прикосновениях и при перемещении. Эти окончания могут служить как полимодальные неспецифические болевые рецепторы только при патологических условиях, например, вследствие возрастания их чувствительности (сенсибилизации) медиаторами воспаления. Слабое раздражение полимодальных неспецифических тактильных рецепторов приводит к чувству зуда. Порог их возбудимости понижают гистамин и серотонин .
Специфическими первичными болевыми рецепторами (нонирецепторами) служат два других типа нервных окончаний – тонкие миелинизированные Аδ-терминали и тонкие немиелинизированные С-волокна , филогенетически более примитивны. Оба эти типа терминалей представлены и в поверхностных тканях, и во внутренних органах. Ноцирецепторы дают чувство боли в ответ на самые разные интенсивные стимулы – механическое воздействие, термический сигнал и т.д. Ишемия всегда вызывает боль поскольку провоцирует ацидоз. Мышечный спазм может вызвать раздражение болевых окончаний из-за относительной гипоксии и ишемии, которые он вызывает, а также вследствие прямого механического смещения ноцирецепторов. По С-волокнам проводится со скоростью 0,5-2 м/с медленная, протопатическая боль , а по миелинизированным, быстропроводящим Аδ-волокнам, обеспечивающим скорость проведения от 6 до 30 м/с, - эпикритическая боль . Кроме кожи, где, по данным А.Г.Бухтиярова, насчитывается не менее 100-200 болевых рецепторов на 1 см, слизистых и роговицы, болевыми рецепторами обоих типов обильно снабжены надкостница, а так же сосудистые стенки, суставы, мозговые синусы и париетальные листки серозных оболочек. В висцеральных листках этих оболочек и внутренних органов болевых рецепторов гораздо меньше.
Боли при нейрохирургических операциях максимальны в момент рассечения мозговых оболочек, в то же время кора больших полушарий обладает очень незначительной и строго локальной болевой чувствительностью. Вообще такой распространенный симптом как головная боль, практически всегда связан с раздражением болевых рецепторов вне самой ткани мозга. Экстракраниальной причиной головной боли могут быть процессы локализованные в синусах костей головы, спазм цилиарной и других глазных мышц, тоническое напряжение мышц шеи и скальпа. Интракраниальные причины головной боли – это в первую очередь раздражение ноцирецепторов мозговых оболочек. При менингите сильнейшие головные боли охватывают всю голову. Весьма серьезную головную боль вызывает раздражение ноцирецепторов в мозговых синусах и артериях, особенно в бассейне средней мозговой артерии. Даже незначительные потери цереброспинальной жидкости могут спровоцировать головную боль, особенно, в вертикальном положении тела, поскольку плавучесть мозга меняется, и при уменьшении гидравлической подушки раздражаются болевые рецепторы его оболочек. С другой стороны, избыток цереброспинальной жидкости и нарушение ее оттока при гидроцефалии, отек головного мозга, его набухание при внутриклеточной гипергидратации, полнокровие сосудов мозговых оболочек, вызванное цитокинами при инфекциях, локальные объемные процессы – также провоцируют головную боль, т.к. при этом увеличивается механическое воздействие на болевые рецепторы окружающих собственно мозг структур.
Болевые рецепторы претендуют на уникальное положение в человеческом теле. Это единственный тип чувствительных рецепторов, которые не подлежат какой бы то ни было адаптации или десенсибилизации под воздействием длящегося или повторяющегося сигнала. Ноцирецепторы при этом не превышают порог своей возбудимости, подобно, например, холодовым сенсорам. Следовательно, рецептор не «привыкает» к боли. Более того, в ноцирецептивных нервных окончаниях имеет место прямо противоположное явление – сенсибилизация болевых рецепторов сигналом . При воспалении, повреждениях ткани и при повторных и длительных болевых раздражителях порог болевой возбудимости ноцирецепторов снижается. Называя болевые сенсоры рецепторами необходимо подчеркнуть, что применение к ним этого термина носит условный характер – ведь это свободные нервные окончания, лишенные каких бы то ни было специальных рецепторных приспособлений.
Нейрохимические механизмы раздражения ноцирецепторов хорошо изучены. Их основным стимулятором является брадикинин . В ответ на повреждение клеток близ ноцирецептора освобождаются этот медиатор, а так же простагландины, лейкотриены, иона калия и водорода . Простагландины и лейкотриены сенсибилизируют ноцирецепторы к кининам, а калий и водород облегчает их деполяризацию и возникновения в них электрического афферентного болевого сигнала. Возбуждение распространяется не только афферентно, но и антидромно, в соседние ветви терминали. Там оно приводит к секреции вещества Р . Этот нейропептид вызывает вокруг терминали паракринным путем гиперемию, отек, дегрануляцию тучных клеток и тромбоцитов. Освобождаемые при этом гистамин , серотонин, простагландины сенсибилизируют ноцирецепторы, а химаза и триптаза мастоцитов усиливают продукцию их прямого агониста – брадикинина. Следовательно, при повреждении ноцирецепторы действуют как сенсоры, и как паракринные провокаторы воспаления. Вблизи ноцирецепторов, как правило, располагаются симпатические норадренергические постганглионарные нервные окончания, которые способны модулировать чувствительность ноцирецепторов .
При травмах периферических нервов нередко развивается так называемая каузалгия – патологически повышенная чувствительность ноцирецепторов в области, иннервируемой поврежденным нервом , сопровождаемая жгучими болямии доже признаками воспаления без видимых местных повреждений. Механизм каузалгии связан с гипералгизирующим действием симпатических нервов, в частности, выделяемого ими норадненалина, на состояние болевых рецепторов. Возможно, при этом происходит секреция вещества Р и других нейропептидов симпатическими нервами, что и обуславливает воспалительные симптомы .
5.2. Система эндогенной модуляции боли.
В контроле возбудимости нейронов, передающих в ЦНС болевые импульсы, принимают участие в основном опиатэргические, серотонинэргические и норадренэргические воздействия. Анатомически, структурами, где сосредоточены элементы модулирующей систеиы являются таламус, серое вещество в окружности сильвиева водопровода, ядра шва, гелеподобное вещество спинного мозга и nucleus traсtus solitarii .
Входные сигналы от лобной коры и гипоталамуса могут активировать энкефалинэргические нейроны вокруг водопровода Сильвия, в среднем мозге и мосте. От них возбуждение нисходит на большое ядро шва, пронизывающее нижнюю часть моста и верхнюю – продолговатого мозга. Нейротрансмиттером в нейронах этого ядра является серотонин . Антиболевой центральный эффект серотонина связан с его антидепрессивным и противотревожным действием .
Ядро шва и близкие к нему роствентрикулярные нейроны продолговатого мозга проводят антиноцирецептивные сигналы в задние рога спинного мозга, где их воспринимают энкефалинэргические нейроны substantia grisea. Энкефалин, вырабатываемый этими тормозными нейронами, осуществляет пресинаптическое ингибирование на болевых афферентных волокнах. Т.о., энкефалин и серотонин передают друг другу эстафетную палочку противоболевой сигнализации . Именно поэтому, морфин и его аналоги, а также агонисты и блокаторы захвата серотонина заняли важное место в анестезиологии. Блокируются не только оба типа болевой чувствительности. Торможение распространяется на защитные болевые спинальные рефлексы, осуществляется оно и на супраспинальном уровне. Опиатэргические системы тормозят стрессорную активности в гипоталамусе (здесь наиболее важен β-эндорфин), ингибируют активность центров гнева, активируют центр наград, вызывают через лимбическую систему изменение эмоционального фона, подавляя отрицательные болевые эмоциональные корреляты и понижают активирующее действие боли на все отделы ЦНС .
Эндогенные опиоиды через спинномозговую жидкость могут попасть в системный кровоток для осуществления эндокринной регуляции, подавляющей системные реакции на боль.
Все способы распространения нейропептидов составляют так называемый трансвентрикулярный путь гипоталамической регуляции.
Депрессии, сопровождаемые уменьшение продукции опиатов и серотонина, часто характеризуются обострением болевой чувствительности . Энкефалины и холецистокинин являются пептидными ко-трансмиттерами в дофаминэргических нейронах. Хорошо известно, что дофаминэргинческая гиперактивность в лимбической системе является одной из патогенетических особенностей шизофрении .